
The introduction to this series can be found here.
Summary: Genetic evidence shows that various Pleistocene populations interbred, including humans, Neanderthals and Denisovans. Is it possible to know whether these could have occurred in pair-bonded relationships? What were the evolutionary origins of pair-bonds in hominins, or are single explanations too simple?
____________________________________________________________________
.
Paleoanthropologist John Hawks created this great infographic that summarizes what we know about ancient human interbreeding, based on recent genetic discoveries (this link to his site has a larger version). The graphic shows that several archaic populations mated with each other, although likely at low rates, including modern humans with Neanderthals and Denisovans. There are also a couple of mystery populations in the mix, whose existence is known solely based on the DNA they left behind. Very cool stuff.
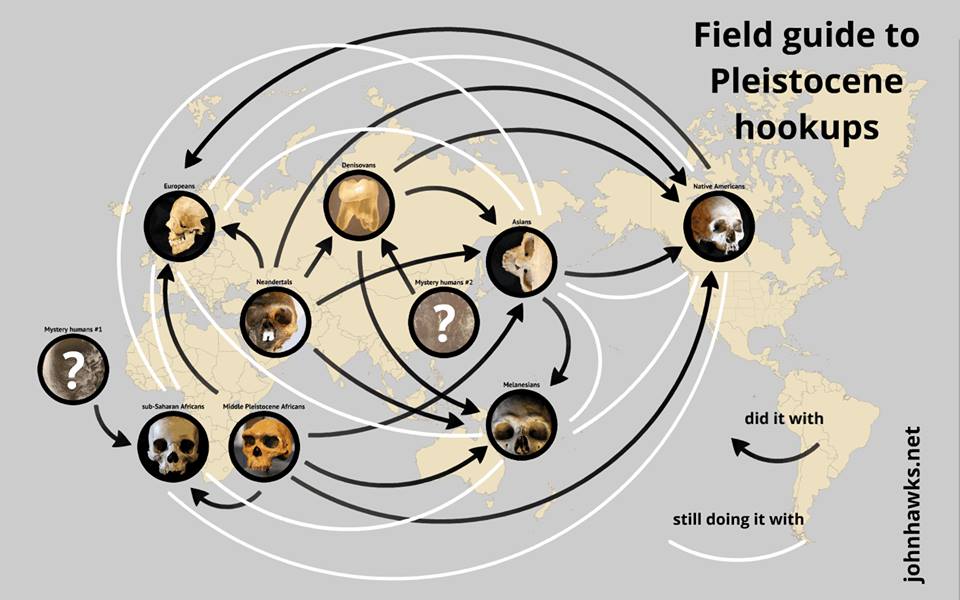
Who Interbred with Whom in the Pleistocene. (Figure by John Hawks).
.
This passage from the Nature paper by Kay Prüfer et al. (2014) gives a nice summary:
“An analysis of the relationships and population history of available archaic genomes and 25 present-day human genomes shows that several gene flow events occurred among Neanderthals, Denisovans and early modern humans, possibly including gene flow into Denisovans from an unknown archaic group. Thus, interbreeding, albeit of low magnitude, occurred among many hominin groups in the Late Pleistocene.”
A few thoughts:
1. The amount of interbreeding (gene flow) may have been low in magnitude, but the fact that gene flow was even possible essentially means that, as Hawks succinctly put it: “Ancient human populations like the Neandertals and Denisovans were not separate biological species.” In fact, Holly Dunsworth asks us to reconsider the concept of Neanderthal extinction altogether. After all, she says, if she is genetically 2.9% Neanderthal, are they really extinct if those genes are still with us today? Personally, I’m only 2.7%, but what this means is that I have some Neanderthals in my family tree, as do most people with sizable ancestry outside of Africa.
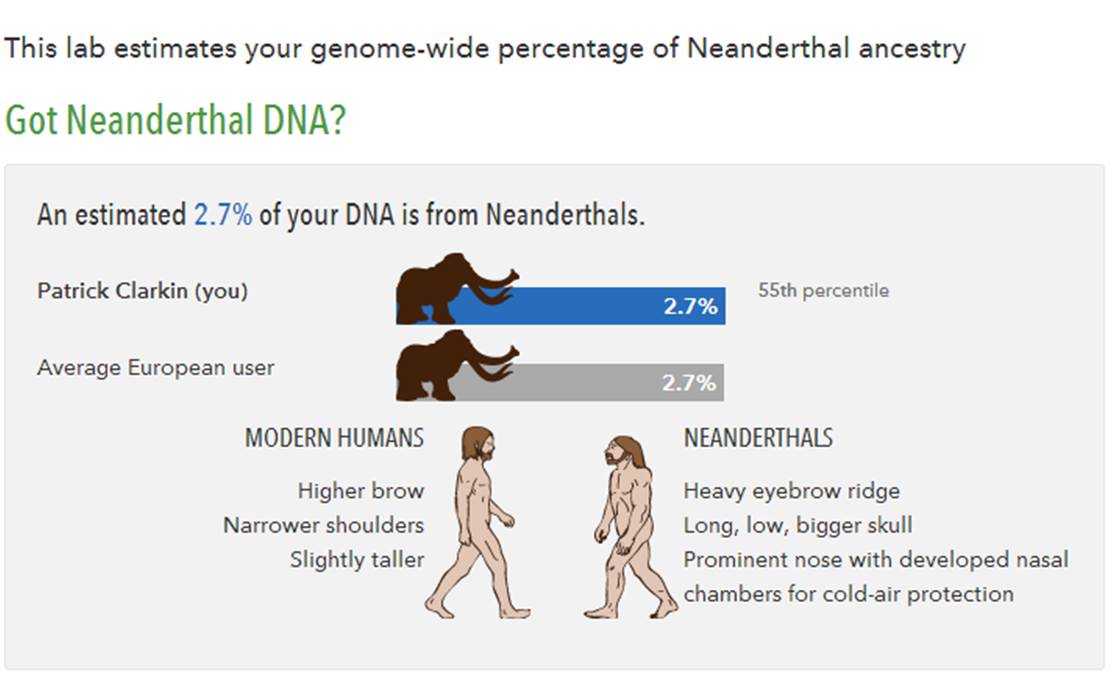
55th percentile. Pretty average.
2. These findings help confirm that gene flow among diverse human groups is not a recent phenomenon. It’s likely that whenever opportunities presented themselves, archaic human populations mated with their neighbors, going back tens of thousands of years ago (at least). It’s telling that a synonym for gene flow is simply ‘migration.’ That’s what humans, archaic and modern, do when they migrate: they mate with who’s around. Whatever barriers existed for them — differences in language, physical appearance, culture, etc. — they were not insurmountable.
3. Estimates of interbreeding rates are not equal to contact rates or rates of sexual liaisons. Finding Neanderthal or Denisovan DNA in other lineages is evidence of matings that resulted in fertile offspring whose genes were passed down for generations, even to the present. It’s a mathematical given that there were many more liaisons that did not have the same fortuitous outcomes, genetically speaking, where pregnancy did not result or those offspring did not themselves reproduce somewhere along the way. In any case, those liaisons may not have been everyday occurrences, but neither were they rare or isolated.
Speculations About Archaic Love
These discoveries about interbreeding are real and important, but with tongue-in-cheek Hawks describes them as ‘Pleistocene hookups’ (he did categorize his post under ‘humor’). The character of these encounters is necessarily speculative. Without getting too prurient, we don’t know if they were brief encounters, or if they could have occurred within pair-bonds. Were these archaic ‘lovers’? (Ugh, that word bums me out). It’s also possible that at least some of these encounters were forcible, as disturbing as that suggestion is. All we have are the genetic outcomes. Some, like National Geographic, have played up the racier view of what might have happened when humans and Neanderthals met up.

.
The truth is that we’ll probably never really know exactly how this all played out, but we can make some educated guesses. We have to try, right? Understanding how pair-bonding originated, and how this might impact modern lives is kind of important stuff. Robert Martin (2013), in his book How We Do It, wrote that the complexity of the question is enough to make some people want to throw their hands up in the air and just give up (but we can’t):
“Which brings us to a fundamental evolutionary question: Are humans biologically adapted for a particular pattern of social organization and mating? Cross-cultural evidence indicates that, as a species, we are highly variable in both respects. Evidence from comparisons with nonhuman primates is equally inconclusive. Despite this ambiguity, many authors resort to a shortcut. They simply take the chimpanzee as a frozen ancestor for human origins, concluding that our evolution began with promiscuous, multimale groups. At the other extreme, it is reasonable to argue that available evidence is so weak that we really cannot reach any firm conclusion regarding our ancestry. Indeed, many would conclude that there is no biological basis whatsoever for human social organization or mating arrangements and that social convention governs everything. According to this view, monogamous marriage is a purely social construct free of any biological predisposition. It turns out that both extreme views are untenable.” (p. 62-3)
OK, so how can we attempt to figure all this out?
1. A number of anthropologists have proposed that pair-bonds, and possibly romantic love, could have ancient origins. Helen Fisher speculated that the earliest neurobiological changes involved in romantic love in hominins could date back to 4.4 million years ago (greatly predating modern humans, Neanderthals, and Denisovans), and going back to some of the earliest species in our lineage. Whatever we call it, romantic love is a real biological phenomenon (Part 5), albeit complex (Part 9). The fact that it is found in nearly all modern human cultures surveyed is is a very strong hint that this dates at least to the beginning of our species. Could it have evolved earlier? If so, by how much? These are tough questions.
Fisher’s claim stems from Owen Lovejoy’s (2009) observation that at 4.4 mya, Ardipithecus ramidus showed reduced differences in size (low sexual dimorphism) between males and females, and smaller canine teeth. From there, he inferred that smaller canines in males could have meant less male-male aggression and less competition for mates. This implied, in turn, relatively long-term social and sexual pair-bonds, where males would have engaged in ‘targeted provisioning’ by bringing food to their preferred mate and their offspring. Ergo, according to Helen Fisher, the beginnings of romantic love.
Recall that pair-bonds are not necessarily equivalent to romantic love or to monogamy, so we aren’t talking about marriages or some Paleolithic version of The Notebook. Fuentes (2012: 188) described pair-bonds somewhat loosely as “a special, predictable relationship between two adults.” Pair-bonds entail a strong psychological affinity between the two, with physiological and emotional responses that differ from other relationships. Sexual pair-bonds include these components, as well as a strong preference to mate with each other compared to other options.
2. On the other hand, 4.4 million years is a long time, and there are so many pieces to this puzzle. It’s very possible that the biology of pair-bonding has some antecedents that predate our species. However, the farther back in time we go, the fuzzier the picture becomes. Another problem, as Robin Dunbar and Susan Shultz (2010) pointed out, is that it’s one thing to look at a species’ behavior and with whom individuals bond and prefer spending their time. But it’s quite another thing to get at the cognitive and emotional processes underlying those bonds. After all, we often have a hard time articulating the complex emotions and drives going on within ourselves. And we spend a lot of our time with ourselves (all of it, actually), so that should be the subject we know best. Trying to understand the internal mental and emotional states within deceased individuals who lived thousands to millions of years ago can only be that much more difficult.
Still, perhaps we can get at some of this indirectly.
3. Duda and Zrzavý (2013) systematically reconstructed how the last common ancestor of modern humans and Pan (bonobos and chimps) might have behaved roughly 6 million years ago, based on 65 life history traits across all living ape species. They concluded that we can be pretty confident about certain features at that stage of evolution, such as that our ancestors: had a primarily single male/multi-female mating system (polygyny), with males being ‘possessive,’ moderate sexual dimorphism, some paternal care, and medium-sized testes. These are not set in stone, and they acknowledge that all primate species show “substantial behavioral diversity and plasticity at both individual and population levels” (p. 429). Still, this is the big picture as they see it.
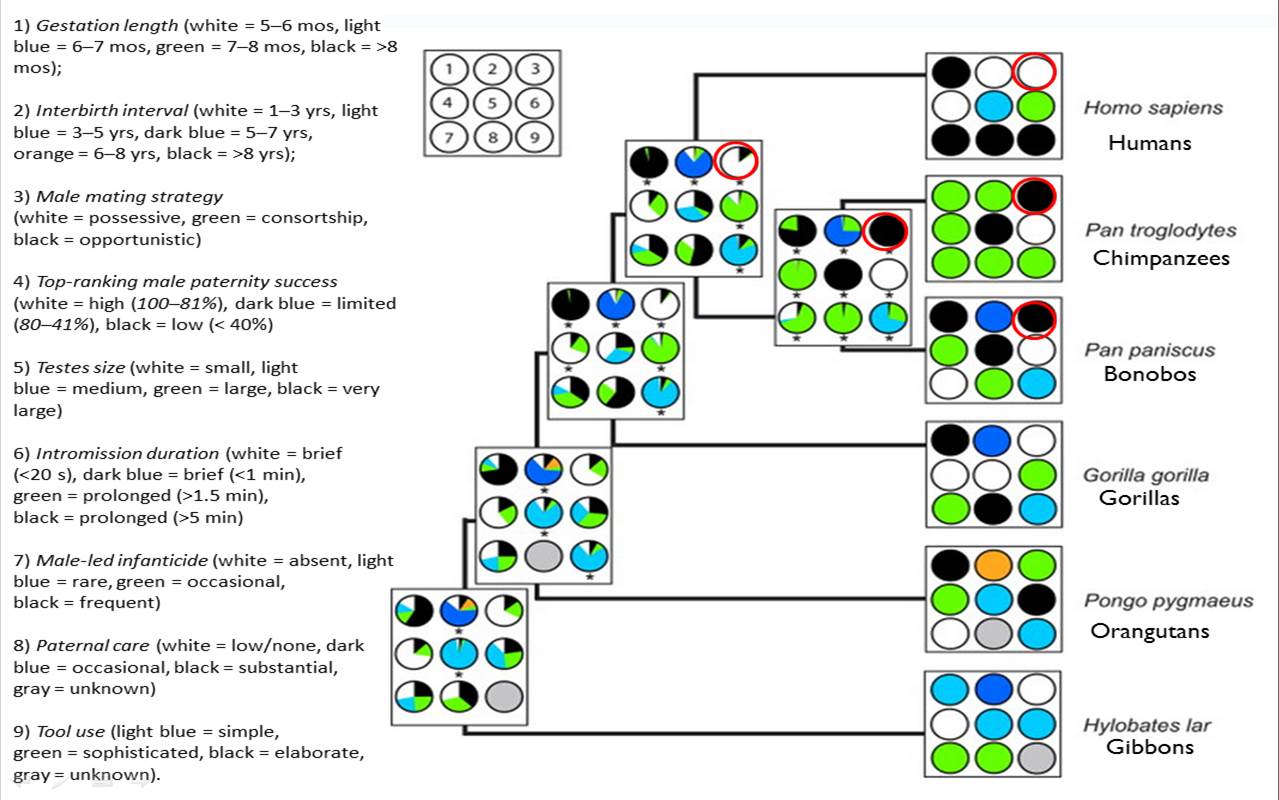
Duda and Zrzavy’s (2013) reconstruction of last common ancestors of humans and apes for 9 traits.
At first glance the above chart is intimidating, but looking at the circles in position three (the top right, a few of which I circled in red), male mating strategies are colored in black, and described as ‘opportunistic’ or uncommitted for chimps, bonobos, and their last common ancestor. This would account for their high sperm counts and large testes, under conditions where there is high competition among males (Part 4). However, unlike the “frozen ancestor” that model Robert Martin warned us about, these traits seem to be derived and evolved after they split from us, rather than something that was present at the time of our last shared ancestor. In other words, this suggests that extreme promiscuity is something chimps and bonobos were moving toward, rather than something we were moving away from.
By contrast, humans are described as being ‘possessive’ (that is, they have enduring mate preferences and engage in mate guarding). Also, this possessiveness was likely present before the split from the chimp line. Therefore, mate guarding in some form likely has been with us for the last few million years, into the various hominin species that followed: Ardipithecus and Neanderthals included. How sure am I about all this? Not very. But it’s something to work with.
Was this equivalent to ‘romantic love,’ as Helen Fisher said? It seems like a pretty big leap to suggest that the neurobiological precursors to romantic love began that early in evolutionary time. Extraordinary claims require extraordinary evidence, and I don’t know if relying on canine teeth from 4.4 mya is enough. Bonds can be maintained through other means. For example, gibbons live in (mostly) monogamous pairs, but do so without exhibiting romantic love.
Others have proposed more moderate dates, with pair-bonding becoming more common around the beginning of our genus, Homo, perhaps around 2 million years ago (ex. see the excellent book by Gray and Garcia, 2013). Alan Dixson (2009: 180) portrayed this transition, with an emphasis on the advantages of bi-parental care for big-brained, slow growing infants and children:
“The propensity for men and women to form long-lasting relationships for reproductive purposes is an ancient trait, probably present in the earliest members of the genus Homo. The occurrence of such relations is universal across extant human populations, and its existence in ancestral forms provided the biological foundation for the later emergence of marriage. Lengthy periods of human infancy, childhood, and adolescence require considerable parental investment by both sexes if offspring are to survive… Nowadays, as throughout recorded history, marriages might be either polygynous or monogamous, depending on the culture considered… During evolution of the genus Homo, a shift towards paired relationships and monogamy has occurred.”
The scenario of pair-bonding evolving for purposes of bi-parental care is fairly accepted, but this is not all encompassing. Quinlan (2008: 235) emphasized that pair-bonds could have had multiple benefits, depending on circumstances. Certainly, more parental care for relatively helpless infants and children would be better than less care. However, pair-bonds might also ensure cooperation and mating stability, particularly when living in large groups. It depends:
“Recent debate about the function of human pair-bonds focuses on mutually exclusive views of men’s work. Thus, men’s work is either for provisioning or for mating effort. This polemic neglects phenotypic plasticity as the key feature of human adaptation to a range of environments.”
Another shortcoming of the bi-parental care scenario is that fathers are not always part of the child-rearing picture. Sarah Hrdy (2009) pointed out how incredibly varied and flexible human fathering is. In hunter-gatherer populations, the amount of paternal investment in childcare is at least partially dependent on circumstances, including the availability of others, such as the mother’s kin, to act as “alloparents.” In support of this, Katie Hinde and colleagues constructed a mathematical model which found that broad cooperative networks, including cooperation among adult females, were a better fit for raising young in early Homo than were male-female pair-bonds alone (see Bell et al 2013).
4. Nelson et al (2011) tried a different approach, looking at finger length ratios in several fossil species (including Ardipithecus, Australopithecus afarensis, and 5 Neanderthals), and then compared them to living species. The rationale behind this is that the ratio of the second-to-fourth digits (2D:4D) acts as a marker for prenatal exposure to sex hormones, with lower ratios associated with higher prenatal androgens (ex. testosterone). Across other primates, the 2D:4D ratio is higher in pair-bonded species compared to more promiscuous or competitive ones.
They found that Ardipithecus was more consistent with non-pairbonded species, contradicting Lovejoy’s and Fisher’s view. Australopithecus was in the pair-bonded range (though it fell well short of values seen in monogamous gibbons). However, the authors were hesitant about the quality of preservation the Australopithecus bones, which came from a single individual.
The picture for the 5 Neanderthals was a bit muddled. The three with the highest ratios (Shanidar 4, Kebara 2, Le Ferrassie I) fell into a pair-bonded or ‘intermediate’ range, while the other two (Le Regourdou and Spy II) were classified as non-pair-bonded (a list of Neanderthal sites can be found here). As Nelson et al put it: “variance in Neanderthal digit ratios might indicate that these Middle and Late Pleistocene hominins, like contemporary humans, exhibited some flexibility in their social systems and mating behaviour across populations.” So much for a definitive answer.
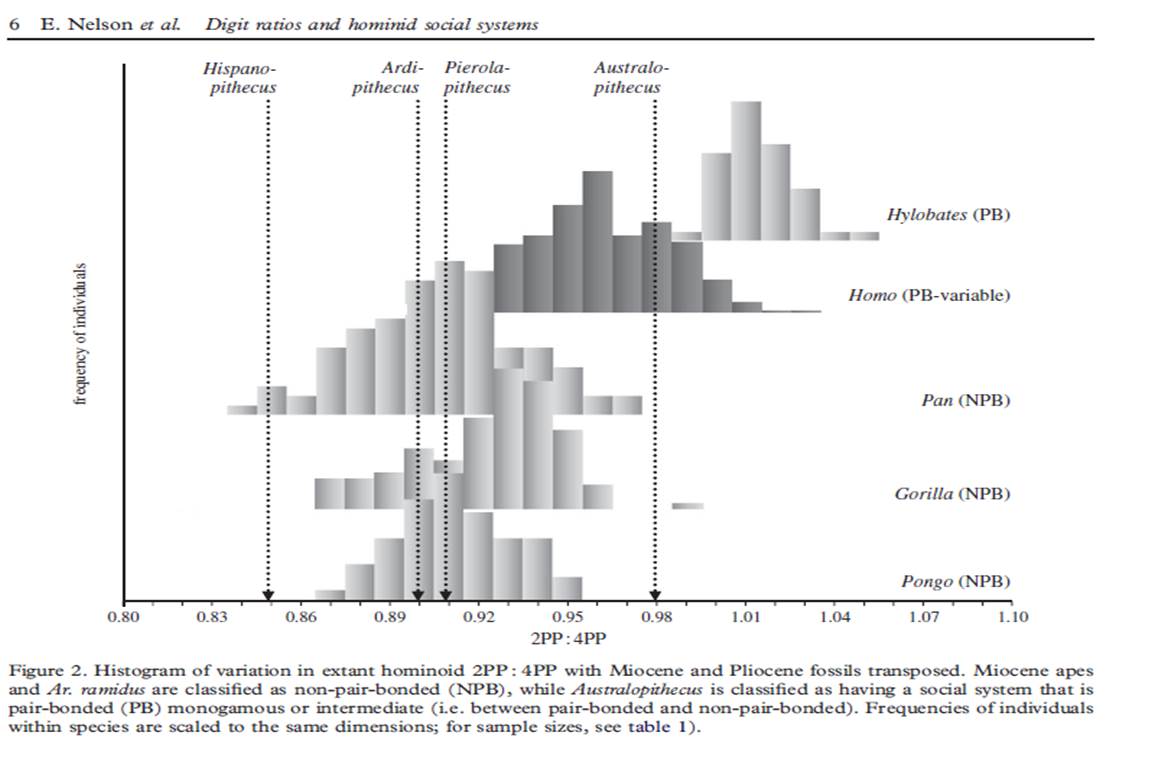
From Nelson et al. Digit ratios of various apes and hominins.
As far as classifying our ancestors’ behavior, I don’t know exactly how much stock to put into finger ratios yet. It would be nice to see more studies like this (where would A. sediba fit?) However, it’s telling that our genus, Homo, is largely intermediate between (with some overlap) the polygynous gorillas, promiscuous chimps, and dispersed orangutans on the one hand, and the monogamous gibbons on the other hand. There we are: not quite monogamous, not quite promiscuous, but as Robert Sapolsky referred to us, “tragically confused”. Another way to look at things is that our genus, including the Neanderthals, has retained flexibility in mating behaviors, allowing them to respond to shifting circumstances.
.
5. Finally, let’s return to the encounters our ancestors had with the Neanderthals and Denisovans. One more thing to consider is what communication would have been like. Dediu and Levinson (2013) argued that speech and language were not acquired suddenly by our species, but rather are much older than previously accepted, perhaps up to half a million years old. D’Anastasio et al. (2013) gave some physical evidence supporting this idea, through the analysis of a Neanderthal hyoid (a bone found in the throat) from Israel. In their view, Neanderthal and modern human hyoids are very similar, suggesting that this is “consistent with a capacity for speech in the Neanderthals.”
If true, and Neanderthals and Denisovans had at least some capacity for language, there could have been some level of communication, and possibly verbalized affection. Of course, even if Neanderthals and Denisovans had the capacity for language, this does not mean that their form of it was equivalent to our ancestors’, and it’s even less likely that they spoke the same language.
Still, these big-brained neighbors on the frontier could have picked up some the traditions and language from each other. Why not? Even if they weren’t fluent in each others’ languages, some form of communication may have taken place. If Augustin Fuentes could communicate with a wild orangutan, in however limited a fashion, then our ancestors could have done so with their fellow hominin neighbors. Could this be relevant to ancient liaisons? Admittedly, this all seems like a longshot, but if we take into account the finger ratios suggesting that Neanderthals were intermediate between being pair-bonded and not, it’s at least in the realm of possibility.
Conclusions
Obviously, trying to put the whole picture together about when and why pair bonding and romantic love gained a toehold in our past is really hard. Our evolutionary story seems to want to defy a simple narrative. One reason for this is that we are not hardwired into having a single strategy. Instead, as Ken Weiss wrote: “we are hard-wired not to be hard-wired.” On the one hand, romantic love and pair-bonding are real, powerful, and integral biological phenomena for our species, and perhaps our genus. On the other, these seem to be a part of a greater tapestry of sexual behavior in our lineage, rather than the whole picture. David Schmitt (2005: 274) looked at the number of sexual partners that people desired across 48 countries, concluding that:
“Clearly, the notion that women are designed solely for lifelong pair bonding, and that any deviation from long-term monogamy represents a maladaptive response of our pair-bonding system, is at odds with the prevailing evidence that multiple mating is a relatively common–and in some ways preferred–sexual strategy.”
Interwoven throughout this series has been an attempt to incorporate complexity and messiness, (especially Parts 1, 6, 10, 13), if only because the idea of our species having a single strategy has to ignore a good deal of inconvenient facts. The sexual pair-bond has a very important place in our behavioral repertoire, probably even the primary one. At the same time, we also seem to have retained some flexibility over our evolutionary past.
.
Series
- Part 1. Introduction Link
- Part 2. Promiscuity Link
- Part 3. Promiscuity (Genetics) Link
- Part 4. Promiscuity (Anatomy/Physiology) Link
- Part 5. Pair-Bonding and Romantic Love Link
- Part 6. Many Intimate Relationships Link
- Part 7. Is It Possible to Love Two People? Link
- Part 8. Love and Suffering Link
- Part 9. Love Is an Evolutionary Compromise Link
- Part 10. Wondrously Complex Paleo-Sex Link
- Part 11. Sexaptation: The Many Functions of Sex Link
- Part 12. A Tripartite Conundrum Link
- Part 13. Is Monogamy ‘Natural?’ Link
- Part 14. Paleo Hookups and Archaic Lovers
.
References
Bell AV, Hinde K, Newson L. 2013. Who Was Helping? The Scope for Female Cooperative Breeding inEarly Homo. PLoS ONE 8(12): e83667. Link
D’Anastasio R, Wroe S, Tuniz C, Mancini L, Cesana DT, et al. (2013) Micro-Biomechanics of the Kebara 2 hyoid and its implications for speech in Neanderthals. PLoS ONE 8(12): e82261. doi:10.1371/journal.pone.0082261. Link
Dediu D and Levinson SC (2013) On the antiquity of language: the reinterpretation of Neandertal linguistic capacities and its consequences. Front. Psychol. 4:397. doi: 10.3389/fpsyg.2013.00397 Link
Dixson A. 2009. Sexual Selection and the Origins of Human Mating Systems. Oxford. Link
Duda P, Zrzavý J. 2013. Evolution of life history and behavior in Hominidae: Towards phylogenetic reconstruction of the chimpanzee–human last common ancestor. Journal of Human Evolution 65: 424-46. Link
Dunbar R. 2012. The Science of Love and Betrayal. Faber and Faber Link
Dunbar RIM and Shultz S. 2010. Bondedness and sociality. Behaviour 147: 775-803. Link
Fuentes A. 2012. Race, Monogamy, and Other Lies They Told You: Busting Myths about Human Nature. University of California Press. Link
Gray P and Garcia J .2013. Evolution and Human Sexual Behavior. Harvard: Cambridge MA. Link
Hrdy SB. 2009. Mothers and Others: The Evolutionary Origins of Mutual Understanding. Belknap. Link
Lovejoy, C.O., 2009. Reexamining human origins in light of Ardipithecus ramidus. Science 326, 74-74e8. Link
Martin R. 2013. How We Do It: The Evolution and Future of Human Reproduction. Basic. Link
Nelson E, Rolian C, Cashmore L, Shultz S. 2011. Digit ratios predict polygyny in early apes, Ardipithecus, Neanderthals and early modern humans but not in Australopithecus. Proceedings of the Royal Society B. Biological Sciences. 278 (1711): 1556-63. Link
Prüfer K, Racimo F, Patterson N, Jay F et al. 2014. The complete genome sequence of a Neanderthal from the Altai Mountains. Nature 505, 43–49 doi:10.1038/nature12886 Link
Quinlan RJ. 2008. Human pair-bonds: evolutionary functions, ecological variation, and adaptive development. Evolutionary Anthropology 17:227-38. Link
Schmitt DP. 2005. Sociosexuality from Argentina to Zimbabwe: a 48-nation study of sex, culture, and strategies of human mating. Behavioral and Brain Sciences 28(2):247-75; discussion 275-311. Link
.

I don’t want to come off as making a semantic nitpick or to be attacking Hrdy’s work (which I like quite a lot), but I feel like there are some significant biases bundled up in the term ‘alloparent.’ The whole idea that the parents should necessarily do the primary work of raising their offspring is even bundled into the verb ‘parent.’ I do come from a place where it isn’t uncommon for individuals-other-than-biological-parents to be the ones doing the primary work of raising a child, so maybe the bias is mine.
Hi Matthew, it’s not nitpicking. That’s a good point — parent and alloparent are terms with multiple meanings attached. I find myself wrestling with other related vocabulary too. One example is ‘mate.’ To mate could mean simply ‘to copulate,’ while another is ‘to pair-off with another for some lengthy period of time.’ These are very different things. And a third alternative is the British meaning ‘friend.’ Another great example might be ‘love,’ which I’ve been thinking about a lot. It’s loaded with different connotations – love of children, early romantic love, even love of inanimate objects. Psychologically, these all seem to be different things. Words accumulate baggage, and then we are expected to understand which of the specific meanings a person is using, but it’s not always clear.
A married couple as described in work from Morgan’s era doesn’t mean the couple had a church wedding (plenty of people didn’t back then), but it did mean that they were having sex. And kinship and marriage were typically taken to be two separate, if related, topics.
Anyway, my point being that it’s difficult to get at the earliest kinship literature without some background of what the authors took for granted. I wonder if the same will be true of things written today being read a century from now?
Pingback: Morsels For The Mind – 31/01/2014 › Six Incredible Things Before Breakfast
Nice piece! The archaeological record alone is so hard to interpret for questions of how Neandertals were socially organised, and what mating mechanisms were. Main proxy used for territory size and social exlusivity is stone tool transfer distances, but this is really problematic: I doubt it measures what people have said it does, and to me seems probably much more about wider techno-economic organisation, mobility systems etc.
I think the fact that Neandertal breeding with AMH happened at all is revealing about the former’s capacity for inter-group communication. More aDNA studies from burial sites would be interesting too in terms of seeing what the levels of inbreeding were both among Neandertals and AMH.
Pingback: 2014 in Review | Patrick F. Clarkin, Ph.D.